全國服務咨詢熱線:
15921799099
15921799099


炎癥小體是多蛋白復合物,其活性與生理性和病理性炎癥密切相關。炎癥小體激活的標志性事件為細胞因子白細胞介素-1β(IL-1β)和白細胞介素-18(IL-18)的成熟,以及gasdermin D蛋白的加工——該過程介導固有免疫系統細胞的細胞因子釋放和焦亡。
炎癥小體是一類高分子量復合物,可激活炎性半胱天冬酶(caspase)和白細胞介素-1家族細胞因子(IL-1β、IL-18,受刺激不同還可能包括IL-1α)。目前已發現多種炎癥小體,其包含的傳感器蛋白各有不同,如NLRP1(NALP1)、NLRP3(NALP3)、IPAF(NLRC4)、NLRP6(NALP6)、NLRP10(NALP10)、NLRP12(NALP12)、Pyrin蛋白、維甲酸誘導基因I(RIG-I)和黑色素瘤缺失蛋白2(AIM-2)。這類炎癥小體大多需要銜接蛋白Asc(含半胱天冬酶募集結構域的凋亡相關斑點樣蛋白)將半胱天冬酶-1募集至炎癥小體復合物中。半胱天冬酶-1與炎癥小體結合后發生切割并激活,進而切割其多種靶蛋白,促使促炎性細胞因子IL-1β成熟并分泌。
炎癥小體可通過多種信號激活,包括活菌、微生物毒素、異源化合物、顆粒物質、細胞質中的病原體相關分子模式(PAMPs)和/或內源性損傷相關分子模式(DAMPs)。相關細節可參考我司的NLRP3炎癥小體掛圖。炎癥小體的活性與多種炎癥反應的誘導存在因果關系,這些反應對機體既可能有益,也可能有害。
有益的炎癥反應可維持組織的穩態功能(如創傷或病原體入侵后檢測并修復組織損傷);有害的炎癥反應則包括顆粒誘導的無菌性炎癥——如尿酸單鈉(MSU)結晶等宿主來源顆粒引發的炎癥(與痛風發病機制相關),以及石棉、二氧化硅、金屬納米顆粒等環境和工業顆粒經吸入后誘導的肺部炎癥。
越來越多的證據表明,炎癥小體的活性還與多種其他疾病相關,包括癌癥、代謝性疾病(如2型糖尿病、動脈粥樣硬化)、部分神經退行性疾病(如阿爾茨海默病、朊病毒病、帕金森病)、自身免疫性疾病(如多發性硬化癥)和炎癥性腸病。此外,炎癥小體還能提升疫苗效力,對宿主發揮有益作用。
1. NLRP3炎癥小體的激活與調控,J. Xu & G. Nunez,《生物化學與生物物理進展》,2023年,第48卷,331頁
2. 定位與細胞信號的協同作用如何激活NLRP3炎癥小體,A. Akbal等,《細胞與分子免疫學雜志》,2022年,第19卷,1201頁
3. 炎性半胱天冬酶:炎癥小體激活半胱天冬酶的統一模型,C. Ross等,《免疫學年度綜述》,2022年,第40卷,249頁
抗NLRP3/NALP3單克隆抗體(Cryo-2),貨號AG-20B-0014-C100,規格100微克
克隆號 | Cryo-2 |
同型 | 小鼠IgG2b |
免疫原 | 重組小鼠NLRP3/NALP3(吡喃結構域/1-93位氨基酸) |
應用 | 免疫細胞化學(ICC)、免疫組化(IHC)、免疫沉淀(IP)、蛋白質免疫印跡(WB,工作濃度1微克/毫升),詳見在線實驗方案 |
特異性 | 識別人和小鼠的NLRP3/NALP3 |
引用量超1000次! |
|
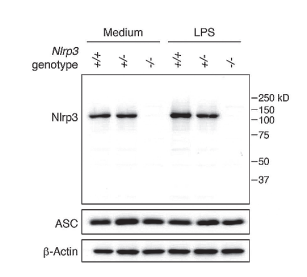
實驗圖:采用NLRP3單克隆抗體(Cryo-2,貨號AG-20B-0014-C100)在小鼠巨噬細胞中檢測到小鼠NLRP3蛋白。
半胱天冬酶-1檢測、標準及其他抗體、預激活與激活、NLRP3抑制劑/激活劑、代謝調控、微管組裝、炎癥小體/焦亡、Gasdermin D與E、鞭毛蛋白與NLRC4、研究進展
●蛋白質免疫印跡(WB)檢測活化的小鼠p10和p20型半胱天冬酶-1
1.抗小鼠半胱天冬酶-1(p10)單克隆抗體(Casper-2),貨號AG-20B-0044-C100(100微克)、AG-20B-0044B-C100(生物素標記,100微克)
克隆號 | Casper-2 |
同型 | 小鼠IgG2a |
免疫原 | 重組小鼠半胱天冬酶-1 |
應用 | 蛋白質免疫印跡(WB,工作濃度1微克/毫升),詳見在線實驗方案 |
特異性 | 識別小鼠內源性全長及活化的半胱天冬酶-1(p10片段) |
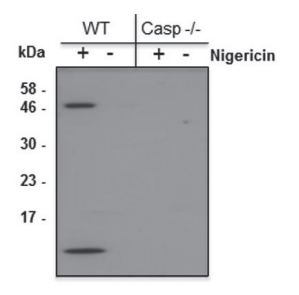
實驗圖:采用抗小鼠半胱天冬酶-1(p10)單克隆抗體(Casper-2,貨號AG-20B-0044),通過免疫印跡在經5微摩爾尼日利亞菌素(貨號AG-CN2-0020)處理30分鐘的野生型和半胱天冬酶-1基因敲除小鼠的分化骨髓源樹突狀細胞(BMDCs)上清液中檢測到小鼠半胱天冬酶-1(p10)。
2.抗小鼠半胱天冬酶-1(p20)單克隆抗體(Casper-1),貨號AG-20B-0042-C100(100微克)、AG-20B-0042B-C100(生物素標記,100微克)
克隆號 | Casper-1 |
同型 | 小鼠IgG1 |
免疫原 | 重組小鼠半胱天冬酶-1 |
應用 | 蛋白質免疫印跡(WB,1微克/毫升)、免疫組化(IHC,石蠟切片)、免疫沉淀(IP),詳見在線實驗方案 |
特異性 | 識別小鼠內源性全長及活化的半胱天冬酶-1(p20片段) |
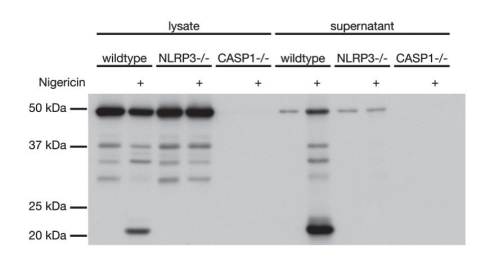
實驗圖:采用抗小鼠半胱天冬酶-1(p20)單克隆抗體(Casper-1,貨號AG-20B-0042),通過免疫印跡在野生型、NLRP3基因敲除和半胱天冬酶-1基因敲除小鼠的分化骨髓源樹突狀細胞(BMDCs)的細胞提取物和上清液中檢測到小鼠半胱天冬酶-1(p20)。
3.抗人半胱天冬酶-1(p20)單克隆抗體(Bally-1),貨號AG-20B-0048-C100(100微克)、AG-20B-0048B-C100(生物素標記,100微克)
克隆號 | Bally-1 |
同型 | 小鼠IgG1 |
免疫原 | 重組人半胱天冬酶-1 |
應用 | 蛋白質免疫印跡(WB,1微克/毫升),詳見在線實驗方案 |
特異性 | 識別人內源性全長及活化的半胱天冬酶-1(p20片段) |

實驗圖:采用抗人半胱天冬酶-1(p20)單克隆抗體(Bally-1,貨號AG-20B-0048),通過免疫印跡檢測到人半胱天冬酶-1(p20)。
實驗方法:用0.5微摩爾佛波酯(PMA,貨號AG-CN2-0010)誘導THP1細胞分化3小時,經5微摩爾尼日利亞菌素處理1小時(激活組,泳道2)或不處理(未激活組,泳道1)后,取上清液通過蛋白質免疫印跡分析半胱天冬酶-1。取30微升上清液經還原條件下的十二烷基-聚丙烯酰胺凝膠電泳(SDS-PAGE)分離,轉印至硝酸纖維素膜,與1微克/毫升的抗人半胱天冬酶-1(p20)單克隆抗體(Bally-1)孵育,采用化學發光檢測系統顯色。
一種替代蛋白質免疫印跡的定量檢測方法,可檢測炎癥小體激活引發的半胱天冬酶-1切割與分泌。
1. 小鼠半胱天冬酶-1配對檢測試劑盒,貨號AG-46B-0003-KI01,規格5塊96孔板
? 特異性:檢測小鼠半胱天冬酶-1(p10和p20結構域)
? 種屬反應性:小鼠
? 靈敏度:100皮克/毫升
? 檢測范圍:0.15納克/毫升~10納克/毫升
? 檢測類型:比色法/夾心ELISA
? 樣本類型:細胞培養上清液
2. 小鼠半胱天冬酶-1酶聯免疫吸附測定(ELISA)試劑盒,貨號AG-45B-0002-KI01,規格96孔
? 特異性:檢測小鼠半胱天冬酶-1(p10和p20結構域)
? 種屬反應性:小鼠
? 靈敏度:33皮克/毫升
? 檢測范圍:15皮克/毫升~1000皮克/毫升
? 檢測類型:比色法/夾心ELISA
? 樣本類型:細胞培養上清液、血清、血漿
實驗技術縮寫:FACS=流式細胞術;FUNC=功能學應用;ICC=免疫細胞化學;IHC=免疫組化;IP=免疫沉淀;WB=蛋白質免疫印跡
配方縮寫:PF=無防腐劑
種屬縮寫:Hu=人;Ms=小鼠;Rt=大鼠;Rb=兔;Prm=靈長類
抗Asc多克隆抗體(AL177),貨號AG-25B-0006-C100(100微克)、AG-25B-0006PF-C100(無防腐劑,100微克)、AG-25B-0006TS-C100(ATTO 647N標記,100微克)
• 來源:兔
• 免疫原:人Asc蛋白N端對應氨基酸合成肽
• 應用:ICC、IHC(石蠟切片)、IP、WB、功能學應用(抑制)*
• 特異性:識別人和小鼠的Asc蛋白
*可抑制Asc與NLRP3的相互作用,從而在細胞提取物中阻斷半胱天冬酶-1的加工。
Asc抗體(AL177)封閉肽,貨號AG-37B-0001-C100,規格100微克;
Asc(AL177)抗體+封閉肽套裝,貨號AG-44B-2000-KI01,規格1套。
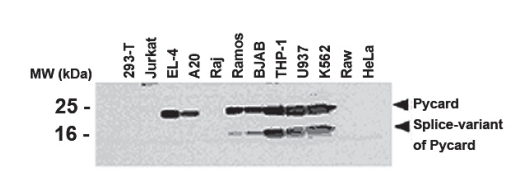
實驗圖:采用抗Asc多克隆抗體(AL177,貨號AG-25B-0006)對人和小鼠細胞系進行蛋白質免疫印跡分析。提取多種人細胞系(293-T、Jurkat、Raj、Ramos、BJAB、THP-1、U937、K562、Raw、HeLa)和小鼠細胞系(EL-4、A20)的總蛋白,經SDS-PAGE分離后,用1:1000稀釋的抗Asc多克隆抗體(AL177)檢測Pycard蛋白,用1:5000稀釋的辣根過氧化物酶偶聯抗兔IgG抗體進行化學發光(ECL)檢測。
分子量(道爾頓):25kDa-Pycard;16kDa-Pycard剪接變體。
抗小鼠NLRP1b單克隆抗體(2A12),貨號AG-20B-0084-C100,規格100微克,應用:WB。
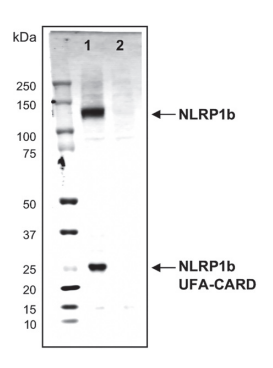
實驗圖:采用NLRP1b單克隆抗體(2A12,貨號AG-20B-0084),在小鼠Raw264.7細胞(泳道1)和NLRP1b基因敲除的Raw264.7細胞(泳道2)的細胞提取物(約30微克)中檢測到小鼠NLRP1b(全長和UFA-CARD片段)。
相關文獻:功能性降解:病原體多種酶激活NLRP1炎癥小體的機制,A. Sandstrom等,《科學》,2019年,第364卷,eaau1330。
抗ZBP1單克隆抗體(Zippy-1),貨號AG-20B-0010-C100,規格100微克,應用:ICC、IP、WB。
雙鏈RNA結合蛋白ZBP1是干擾素生物學中的關鍵因子,參與病毒感染和炎癥調控,可激活NLRP3炎癥小體、干擾素通路,以及由PANoptosome復合物調控的多種程序性細胞死亡。近期研究表明,RNA編輯酶ADAR1可抑制ZBP1的激活。
1. 檢測細菌感染誘導的炎癥小體激活,P. Broz & D.M. Monack,《分子生物學方法》,2013年,第1040卷,65頁
2. 檢測NLR寡聚化Ⅱ:共聚焦顯微鏡和免疫熒光法檢測ASC斑點形成,M. Beilharz等,《分子生物學方法》,2016年,第1417卷,145頁
3. 炎癥小體激活的無細胞實驗,Y. Jamilloux & F. Martinon,《分子生物學方法》,2016年,第1417卷,207頁
4. 流式細胞術檢測人外周血中ASC斑點——炎癥小體激活的讀數指標,N. Wittmann等,《細胞》,2021年,第10卷,2880頁
5. 炎癥小體激活后人原代角質形成細胞中的ASC斑點形成,N. Smatlik等,《氧化醫學與細胞長壽》,2021年,7914829
●Casper-1、Casper-2、Bally-1和Cryo-2相關實驗方案
1. 炎癥小體的檢測,O. Gross,《分子生物學方法》,2012年,第844卷,199頁
2. 活化半胱天冬酶-1的免疫印跡檢測,C. Jakobs等,《分子生物學方法》,2013年,第1040卷,103頁
3. 檢測NLR寡聚化Ⅰ:體積排阻色譜、免疫共沉淀和交聯法,S. Khare等,《分子生物學方法》,2016年,第1417卷,131頁
4. 半胱天冬酶-1激活的檢測,B. Guey & V. Petrilli,《分子生物學方法》,2016年,第1417卷,197頁
5. 炎癥小體激活的無細胞實驗,Y. Jamilloux & F. Martinon,《分子生物學方法》,2016年,第1417卷,207頁
產品名稱 | 貨號 | 規格 | 來源/同型 | 種屬 | 應用 |
NOD樣受體(NLRs) | - | - | - | - | - |
抗小鼠NAIP1/2/5單克隆抗體(Naipa-1) | AG-20B-0045 | 100μg | 小鼠IgG2b κ | 小鼠 | WB |
抗人NLRP1/NALP1多克隆抗體(AL176) | AG-25B-0005 | 100μg | 兔 | 人 | WB |
抗NLRP3/NALP3單克隆抗體(Cryo-2) | AG-20B-0014 | 100μg | 小鼠IgG2b | 人、小鼠 | ICC、IHC、IP、WB |
抗小鼠NLRP3/NALP3單克隆抗體(Cryo-1) | AG-20B-0006 | 100μg | 小鼠IgG2b | 小鼠 | WB |
抗人NLRP6/NALP6單克隆抗體(Clint-1) | AG-20B-0046 | 100μg | 小鼠IgG1 κ | 人 | WB |
RIG樣解旋酶(RLHs)——抗病毒信號通路 | - | - | - | - | - |
抗RIG-I單克隆抗體(Alme-1) | AG-20B-0009 | 100μg | 小鼠IgG1 | 人、小鼠 | IHC、IP、WB |
抗RIG-I單克隆抗體(Alme-1,生物素標記) | AG-20B-0009B | 100μg | 小鼠IgG1 | 人、小鼠 | IHC、IP、WB |
抗人Cardif單克隆抗體(Adri-1) | AG-20B-0004 | 100μg | 小鼠IgG2b | 人 | ICC、IHC、IP、WB |
抗人MDA5單克隆抗體(Hely-1) | AG-20B-0013 | 100μg | 小鼠IgG1 | 人 | ELISA、IP、WB |
抗丙型肝炎病毒NS3單克隆抗體(1B6) | AG-20B-0001 | 100μg | 小鼠IgG1 | 丙型肝炎病毒 | ICC、WB |
抗丙型肝炎病毒NS5B單克隆抗體(5B-3B1) | AG-20B-0002 | 100μg | 小鼠IgG2b | 丙型肝炎病毒 | WB |
抗丙型肝炎病毒NS5B封閉性單克隆抗體(5B-12B7) | AG-20B-0003 | 100μg | 小鼠IgG2a | 丙型肝炎病毒 | ICC、IP、功能學應用(封閉) |
細胞質DNA傳感器 | - | - | - | - | - |
抗人AIM2單克隆抗體(3B10) | AG-20B-0040 | 100μg | 小鼠IgG1 | 人 | ICC、WB |
細胞質細菌傳感器 | - | - | - | - | - |
抗人Pyrin多克隆抗體(AL196) | AG-25B-0020 | 100μg | 兔 | 人 | IP、WB |
細胞質病原體相關分子模式傳感器 | - | - | - | - | - |
抗半胱天冬酶-4/11(p20)單克隆抗體(Flamy-1) | AG-20B-0060 | 100μg | 小鼠IgG2b κ | 人、小鼠 | IP、WB |
抗半胱天冬酶-4/11(p20)單克隆抗體(Flamy-1,生物素標記) | AG-20B-0060B | 100μg | 小鼠IgG2b κ | 人、小鼠 | IP、WB |
七、炎癥小體激活的定量檢測——白細胞介素-1β(IL-1β)
貨號AG-45B-0021-KI01,規格96孔
同型 | 小鼠IgG |
特異性 | 檢測人IL-1β |
種屬反應性 | 人 |
靈敏度 | 0.7皮克/毫升 |
檢測范圍 | 1.5625皮克/毫升~100皮克/毫升 |
檢測類型 | 比色法/夾心ELISA |
樣本類型 | 細胞培養上清液、血清、血漿 |
在病原體相關分子模式(PAMPs)或損傷相關分子模式(DAMPs)等預激活信號作用下,IL-1β以無活性的35kDa前體形式產生;在組織損傷或疾病狀態下,炎癥小體激活后該前體被切割為17kDa的活性形式。活性IL-1β在啟動和擴散無菌性炎癥中發揮已知作用,包括募集巨噬細胞、激活促炎性細胞因子白細胞介素-6(IL-6)以及調控趨化因子的表達。
我司新品人IL-1β ELISA試劑盒可高靈敏度、高特異性地檢測生物體液中的全長和切割型人IL-1β。
抗小鼠IL-1α(p18)單克隆抗體(Teo-1),貨號AG-20B-0064-C100,規格100微克
• 同型:小鼠IgG
• 應用:WB(1微克/毫升)
• 特異性:識別小鼠IL-1α的切割型p18片段和全長片段

實驗圖:采用抗小鼠IL-1α(p18)單克隆抗體(Teo-1,貨號AG-20B-0064),通過免疫印跡檢測到小鼠IL-1α(全長p30片段和切割型p18片段)。
實驗方法:對經脂多糖(LPS)和多種炎癥小體激活劑處理的骨髓源樹突狀細胞(BMDCs)的細胞提取物和上清液進行蛋白質免疫印跡,分析IL-1α。將細胞提取物和上清液經還原條件下的SDS-PAGE分離,轉印至硝酸纖維素膜,與1微克/毫升的抗小鼠IL-1α(p18)單克隆抗體(Teo-1)孵育,加入辣根過氧化物酶(HRP)偶聯的抗小鼠二抗后,采用化學發光檢測系統顯色。
泳道說明:1-脂多糖預激活的骨髓源樹突狀細胞裂解液;2-脂多糖+三磷酸腺苷(ATP)處理的骨髓源樹突狀細胞上清液;3-脂多糖+尼日利亞菌素處理的骨髓源樹突狀細胞上清液;4-脂多糖+尿酸單鈉結晶處理的骨髓源樹突狀細胞上清液。
NLRP3是炎癥小體復合物的核心組分,在炎癥調控中發揮關鍵作用。NLRP3可響應多種刺激,從而抵御多種病毒和細菌感染,如腺病毒、流感病毒、冠狀病毒、金黃色葡萄球菌、鼠傷寒沙門氏菌、單核細胞增生李斯特菌和分枝桿菌。
這些病原體相關分子模式(PAMPs)以及部分非病原性刺激(損傷相關分子模式DAMPs)可通過預激活和激活/觸發兩步過程增強NLRP3炎癥小體的激活。
預激活至少發揮兩大作用:
1. 上調炎癥小體組分NLRP3、半胱天冬酶-1和IL-1β前體的表達:病原體相關分子模式(PAMPs)或損傷相關分子模式(DAMPs)與模式識別受體(PRRs,如Toll樣受體TLRs、核苷酸結合寡聚化結構域蛋白2 NOD2)結合,或腫瘤壞死因子(TNF)、IL-1β等細胞因子激活核因子-κB(NF-κB)并啟動基因轉錄,均可誘導上述組分的轉錄上調;
2. 誘導NLRP3的翻譯后修飾(PTMs):該修飾使NLRP3穩定在自身抑制的無活性但可響應信號的狀態。目前已發現NLRP3的多種翻譯后修飾,包括泛素化、磷酸化和小泛素化修飾(SUMO化)。
與其他模式識別受體不同,NLRP3可被細菌、病毒、真菌感染引發的多種不相關刺激激活,也可在內源性DAMPs介導的無菌性炎癥中,或暴露于環境刺激物后被激活。這些激活劑的共同特征是均能誘導細胞應激,并被NLRP3感知。
NLRP3的激活涉及多種上游信號(多數信號并非互斥),包括鉀離子(K?)/氯離子(Cl?)外流、鈣離子(Ca2?)內流、溶酶體破裂、線粒體功能障礙、代謝變化和反式高爾基體解體。
當病原體相關分子模式(PAMPs)存在或損傷相關分子模式(DAMPs,如ATP、尿酸結晶、活性氧ROS)累積時,NLRP3炎癥小體復合物組裝,進而啟動其實際激活過程。復合物形成后激活半胱天冬酶-1,將IL-1β前體和IL-18前體切割為活性形式,促使這些細胞因子釋放并啟動免疫反應。
此外,翻譯后修飾(PTMs)是NLRP3激活的關鍵調控因子,可影響蛋白質的活性、降解、定位、結構及與其他蛋白的相互作用等多個方面。
1. NLRP3炎癥小體激活調控機制的進展,S. Paik等,《細胞與分子免疫學雜志》,2021年,第18卷,1141頁
2. 炎癥衰老和年齡相關疾病中NLRP3炎癥小體的預激活機制,A. Gritsenko等,《細胞因子與生長因子評論》,2020年,第55卷,15頁
3. NLRP3炎癥小體的預激活:一層套一層的未解之謎,C.M. McKee & R.C. Coll,《白細胞生物學雜志》,2020年,第108卷,937頁
NLRP3炎癥小體最主要的功能是加工并激活IL-1β前體,而多數細胞并不表達IL-1β前體,因此需先誘導其表達。TLR配體、腫瘤壞死因子(TNF)等多種刺激均可誘導IL-1β前體的合成。
脂多糖(LPS) 通過TLR4介導的信號通路發揮作用,因其高效性和易獲取性,始終是誘導IL-1β前體產生的配體;同時,LPS刺激還能觸發NLRP3的多種翻譯后修飾(如泛素化),這是NLRP3預激活過程的關鍵步驟。
1. 佛波醇12-肉豆蔻酸13-乙酸酯(PMA),貨號AG-CN2-0010
2. 可溶性人腫瘤壞死因子-α(重組),貨號AG-40B-0006
3. 多聚體人腫瘤壞死因子-α(重組),貨號AG-40B-0019
4. 多聚體小鼠腫瘤壞死因子-α(重組),貨號AG-40B-0021
5. 脂多糖(LPS)
1. 無菌性炎癥疾病中的炎癥小體預激活,M.N. Patel等,《分子醫學趨勢》,2017年,第23卷,165頁
2. NLRP3激活的關鍵:預激活和溶酶體損傷,V. Hornung & E. Latz,《歐洲免疫學雜志》,2010年,第40卷,620頁
3. 炎癥小體,K. Schr?der & J. Tschopp,《細胞》,2010年,第140卷,821頁
完整產品列表詳見我司《固有免疫研究手冊》。
探索我司多款即用型LPS溶液,專為炎癥小體預激活設計。無需自行溶解LPS,選擇AdipoGen生命科學的均一即用型LPS溶液,直接使用即可。
九、翻譯后修飾(PTMs)和蛋白結合對NLRP3炎癥小體的調控
NLRP3是一種細胞質模式識別受體,在炎癥調控中起核心作用。其預激活和激活受多種翻譯后修飾(PTMs) 調控(如下文),本部分僅為概述,并非涵蓋全部修飾類型。
磷酸化調控NLRP3的活性,是預激活過程的關鍵步驟。NLRP3的磷酸化位點及作用如下:
• 絲氨酸198(小鼠為絲氨酸194):由c-Jun N端激酶1(JNK1)磷酸化,促進NLRP3的去泛素化和寡聚化;
• PYD-NACHT堿性接頭區的4個酪氨酸殘基(Tyr132、Tyr136、Tyr145、Tyr164):由布魯頓酪氨酸激酶(BTK)磷酸化,中和該區域的正電荷,使NLRP3從完整的反式高爾基體網絡(TGN)轉移至分散的反式高爾基體網絡,促進NLRP3炎癥小體組裝;
• 絲氨酸5:由AKT或BTK磷酸化,抑制Asc與NLRP3的結合及炎癥小體激活,該過程可被磷酸酶PP2A逆轉;
• 人絲氨酸295(小鼠絲氨酸293):由蛋白激酶D(PKD)磷酸化,使NLRP3從高爾基體釋放并激活;由蛋白激酶A(PKA)磷酸化,抑制NLRP3的ATP酶活性。
此外,非受體型蛋白酪氨酸磷酸酶22(PTPN22)可與NLRP3相互作用,并使其酪氨酸861位點去磷酸化(該位點的磷酸化激酶尚未明確)。
NLRP3的泛素化和去泛素化調控其穩定性和活性:TLR4和MyD88介導的信號通路可觸發NLRP3的去泛素化,進而誘導炎癥小體激活;去泛素化酶(DUBs)在NLRP3調控中起重要作用,其小分子抑制劑可阻斷NLRP3的激活。
具體調控機制:
• BRCC3介導NLRP3富亮氨酸重復序列(LRR)結構域的去泛素化,是NLRP3寡聚化的必要條件;
• FBXL2使NLRP3的LRR結構域泛素化,促進其經蛋白酶體降解;
• ARIH2與NLRP3的NACHT結構域結合,誘導其K48位連接的泛素化,進而通過蛋白酶體介導降解;
• E3泛素連接酶MARCH7使NLRP3的LRR結構域發生K48位連接的泛素化,通過自噬途徑誘導其降解。
NLRP3的SUMO化調控其穩定性和功能:
• 小泛素樣修飾物(SUMO)E3連接酶MUL1/MAPL可介導NLRP3的SUMO化;NLRP3激活信號可誘導SENP6和SENP7介導其去SUMO化;
• SUMO1介導的SUMO化具有激活作用,是NLRP3激活的必要條件;SENP3介導的去SUMO化則會抑制NLRP3激活;
• 重要發現:LPS預激活對NLRP3的SUMO化無明顯影響。
還包括乙酰化(激活作用,促進NLRP3聚集并與ASC結合)和亞硝基化(NLRP3的負調控因子)。
總體而言,NLRP3、Asc和半胱天冬酶-1的翻譯后修飾在調控NLRP3活性及其炎癥調控功能中發揮關鍵作用。除轉錄和翻譯后調控機制外,多種蛋白結合伴侶也可正向或負向調控NLRP3的預激活和激活,包括:
• NIMA相關激酶7(NEK7,絲氨酸-蘇氨酸激酶):NLRP3炎癥小體組裝和激活的關鍵組分;
• 僅含PYD結構域蛋白(POPs):細胞質中的小型誘餌蛋白,調控炎癥小體激活(僅在人細胞中存在);
• 僅含CARD結構域蛋白(COPs):調控機制尚不明確(僅在人細胞中存在)。
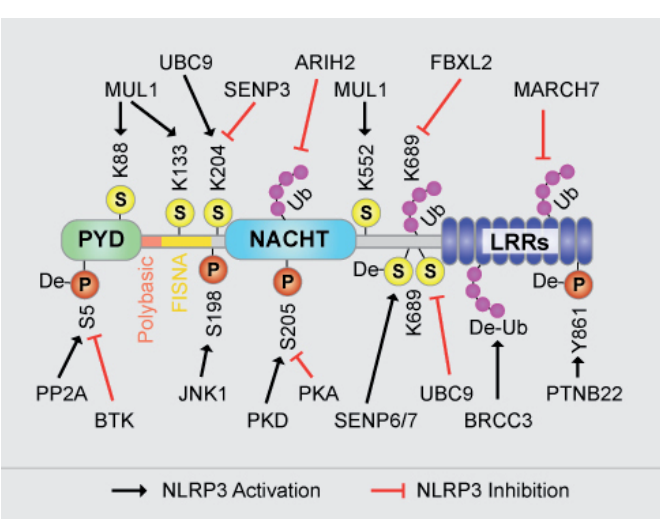
實驗圖:人NLRP3的分子結構與翻譯后修飾(NLRP3激活/抑制相關)。
1. 翻譯后修飾在NLRP3炎癥小體激活調控中的作用,J. Xia等,《國際分子科學雜志》,2023年,第24卷,6126頁
2. 翻譯后修飾對NLRP3炎癥小體的調控,S. Zangiabadi & A.A. Abdul-Sater,《免疫學雜志》,2022年,第208卷,286頁
3. 塑造炎癥小體:磷酸化是炎癥小體信號通路的關鍵開關,C.M. McKee等,《生物化學會匯刊》,2021年,第49卷,2495頁
NLRP3是開發新型抗炎療法的潛力的靶點,NLRP3小分子直接抑制劑已顯示出良好的應用前景。此外,參與NLRP3預激活(轉錄和翻譯后水平)的多種其他蛋白,也可作為靶點以抑制NLRP3介導的炎癥。
產品名稱 | 靶點 | 貨號 |
SP 600125 | JNK抑制劑 | AG-CR1-3549 |
CRT0066101 鹽酸鹽 | PDK抑制劑 | AG-CR1-3550 |
岡田酸 | PP2A抑制劑 | AG-CN2-0056 |
WP1130 | 去泛素化酶(DUB)抑制劑 | AG-CR1-3551 |
β-AP15 | 去泛素化酶(DUB)抑制劑 | AG-CR1-3552 |
貨號:AG-CR1-3950(結晶型,2毫克/2×2毫克/5毫克/25毫克)、AG-CR1-3951(即用型溶液,10毫克/250毫克/1克/5克),經生物活性驗證! 高效NLRP3炎癥小體激活劑。
相關文獻:痛風相關尿酸結晶激活NALP3炎癥小體,F. Martinon等,《自然》,2006年,第440卷,237頁。
●尼日利亞菌素鈉([Nigericin.Na](Nigericin.Na))
貨號AG-CN2-0020,高效NLRP3炎癥小體激活劑。
相關文獻:冷吡啉響應毒素和ATP激活炎癥小體,S. Mariathasan等,《自然》,2006年,第440卷,228頁。
貨號AG-CN2-0489,通過使己糖激酶與線粒體解離,成為NLRP3炎癥小體的新型激活劑。
相關文獻:己糖激酶是檢測細菌肽聚糖的固有免疫受體,A.J. Wolf等,《細胞》,2016年,第166卷,624頁。
產品名稱 | 貨號/規格 | 作用特點 | 相關文獻 |
紫菀內酯(Arglabin) | AG-CN2-0458,1mg/5mg/10mg/50mg | NLRP3炎癥小體抑制劑 | 紫菀內酯的抗炎和抗動脈粥樣硬化作用(高脂飲食ApoE2 Ki小鼠模型),A. Abderrazak等,《循環》,2015年,第131卷,1061頁 |
BAY 11-7082 | AG-CR1-0013,1mg/5mg/10mg/50mg | 抑制NLRP3炎癥小體,降低其ATP酶活性 | 抗炎化合物小白菊內酯和BAY 11-7082是炎癥小體的直接抑制劑,C Juliana等,《生物化學雜志》,2010年,第285卷,9792頁 |
達潘蘇特rile(Dapansutrile) | AG-CR1-3535,10mg/50mg/250mg,可批量供應 | 高效、高選擇性、口服有效的NLRP3抑制劑,直接結合NLRP3 NACHT結構域的ATP結合基序 | - |
MCC950鈉 | AG-CR1-3615,1mg/5mg/10mg,可批量供應 | 高效、高選擇性NLRP3炎癥小體抑制劑 | 1. NLRP3炎癥小體小分子抑制劑用于治療炎癥性疾病,R.C. Coll等,《自然醫學》,2015年,第21卷,248頁;2. NLRP3抑制劑OLT1177抑制小鼠急性關節炎模型的關節炎癥,C Marchetti等,《關節炎研究與治療》,2018年,第20卷,169頁 |
前列腺素E2 | AG-CR1-3613,1g/5g/10g | NLRP3炎癥小體抑制劑 | 前列腺素E2通過EP4受體和細胞內環磷酸腺苷抑制人巨噬細胞的NLRP3炎癥小體激活,M. Sokolowska等,《免疫學雜志》,2015年,第194卷,5472頁 |
格列本脲(USP級) | AG-CL1-0001,1mg/5mg/25mg/100mg | 抑制NLRP3的ATP酶活性(NLRP3-Asc炎癥小體復合物組裝的必要條件) | NLRP3炎癥小體小分子抑制劑用于治療炎癥性疾病,R.C Coll等,《自然醫學》,2015年,第21卷,248頁 |
(S)-3-羥基丁酸/3-羥基丁酸/(R)-3-羥基丁酸 | AG-CR1-3617,25mg/100mg | NLRP3炎癥小體抑制劑,抑制鉀離子外流,進而減少Asc寡聚化和斑點形成 | 酮代謝物β-羥基丁酸阻斷NLRP3炎癥小體介導的炎癥性疾病,Y.H. Youm等,《自然醫學》,2015年,第21卷,263頁 |
小白菊內酯(Parthenolide) | AG-CN2-0455,10mg/50mg/250mg | 抑制NLRP3炎癥小體,降低其ATP酶活性 | 抗炎化合物小白菊內酯和BAY 11-7082是炎癥小體的直接抑制劑,C Juliana等,《生物化學雜志》,2010年,第285卷,9792頁 |
異甘草素(Isoliquiritigenin) | AG-CN2-0459,25mg/100mg | 抑制NLRP3激活的Asc寡聚化,阻斷預激活和激活步驟 | 異甘草素是NLRP3炎癥小體的高效抑制劑,可抑制飲食誘導的脂肪組織炎癥,H. Honda等,《白細胞生物學雜志》,2014年,第96卷,1087頁 |
K777[K11777] | AG-CR1-0158 | 廣譜組織蛋白酶抑制劑,可用于抑制炎癥小體 | 多種組織蛋白酶在NLRP3依賴的IL-1β激活過程中,促進顆粒誘導的非炎癥小體依賴性細胞死亡,GM. Orlowski等,《白細胞生物學雜志》,2017年,第102卷,7頁 |
十三、 炎癥小體及其治療意義
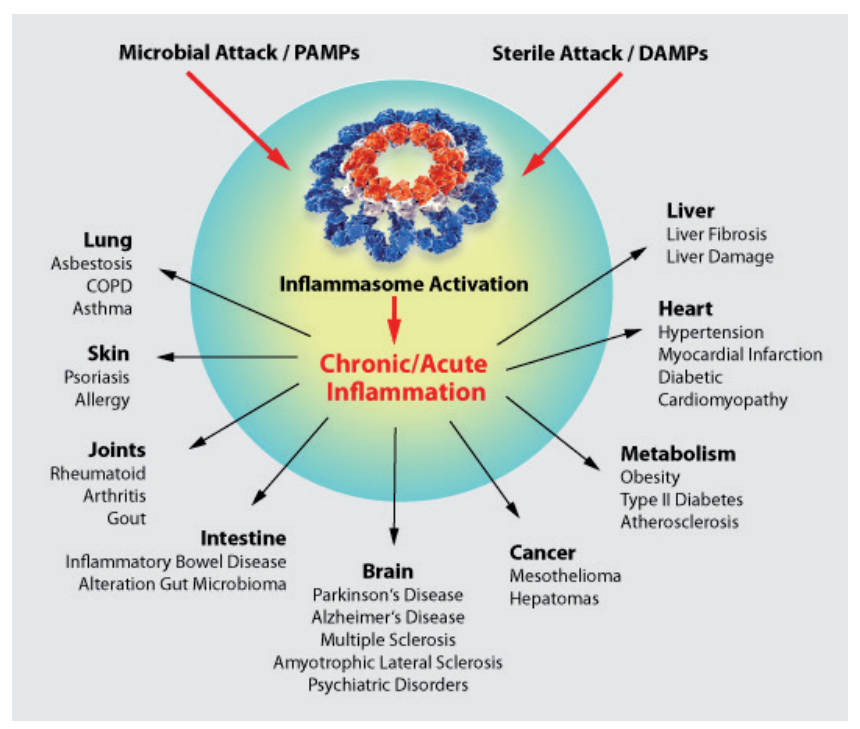
炎癥小體的異常激活與慢性炎癥密切相關,是多種疾病發生發展的誘因——因其在調控免疫反應和促炎性細胞因子產生中發揮關鍵作用。靶向炎癥小體已成為治療多種疾病的潛在策略,包括:
• 炎癥性疾病(如關節炎、痛風、炎癥性腸病);
• 自身免疫性疾病(如紅斑狼瘡、多發性硬化癥);
• 代謝性疾病(如2型糖尿病、肥胖、心血管疾病);
• 癌癥、神經退行性疾病。
近期研究還表明,NLRP3炎癥小體與病毒感染、敗血癥等其他疾病相關。在上述疾病的動物模型中,抑制炎癥小體激活或靶向炎癥小體組分可有效減輕炎癥。總體而言,靶向炎癥小體具有巨大的治療潛力,是開發新型療法的重要方向。
炎癥小體激活相關疾病累及器官/系統:肺(石棉肺、哮喘、慢性阻塞性肺疾病COPD)、皮膚(銀屑病、過敏、急/慢性炎癥)、關節(類風濕關節炎)、代謝系統(肥胖、2型糖尿病)、腸道(炎癥性腸病、腸道微生物群紊亂、痛風)、心臟(高血壓、心肌梗死、糖尿病心肌病)、肝臟(肝損傷、肝纖維化)、大腦(帕金森病、阿爾茨海默病、多發性硬化癥、精神疾病)、其他(癌癥、間皮瘤、肝癌、動脈粥樣硬化)。
1. NLRP3炎癥小體通路抑制劑——炎癥性疾病的潛在治療候選藥物,X. Zhang等,《國際分子醫學雜志》,2023年,第51卷,35頁
2. NLRP3炎癥小體的藥理學抑制:結構、分子激活及抑制劑-NLRP3相互作用,Q. Ma,《藥理學評論》,2023年,第75卷,487頁
3. 聚焦NLRP3炎癥小體在疾病中的作用,R. Fusco等,《國際分子科學雜志》,2020年,第21卷,4223頁
免疫代謝是研究免疫細胞內的代謝過程和通路如何影響其功能的學科。免疫細胞在炎癥反應中被激活時,會發生顯著的代謝變化以滿足其增加的能量需求,主要表現為從氧化磷酸化向糖酵解轉換——糖酵解產能更快但效率更低。
炎癥小體的活性受細胞內代謝通路變化的嚴格調控,近期研究表明,調控免疫代謝可調節免疫反應、降低炎癥小體活性并減輕炎癥。例如,促進免疫細胞的脂肪酸代謝或阻斷其糖酵解,可誘導抗炎反應。
1. NLRP3炎癥小體:代謝信號的調控作用,A. Olona等,《免疫學趨勢》,2022年,第43卷,978頁
2. 靶向免疫代謝作為抗炎策略,E.M. Palsson-McDermott & L. O’Neill,《細胞研究》,2020年,第30卷,300頁
產品名稱 | 靶點 | 貨號 |
2-脫氧-D-葡萄糖 | 己糖激酶抑制劑 | AG-CR1-3681 |
富馬酸二甲酯 | Nrf2激活劑 | AG-CR1-3701 |
庚二酸 | 甘油醛-3-磷酸脫氫酶(GADPH)抑制劑 | AG-CN2-0118 |
鹽酸二甲雙胍 | 活性氧(ROS)抑制劑 | AG-CR1-3689 |
雷帕霉素 | 哺乳動物雷帕霉素靶蛋白(mTOR)抑制劑 | AG-CN2-0025 |
TEPP-46 | 丙酮酸激酶M2(PKM2)激活劑 | AG-CR1-3687 |
微管是細胞骨架的重要組分,除參與細胞分裂、遷移和形態維持等基本功能外,在固有免疫中也發揮關鍵作用。炎癥小體由模式識別受體、銜接蛋白Asc和半胱天冬酶-1組裝而成,可響應病原體相關分子模式(PAMPs)或損傷相關分子模式(DAMPs),加工IL-1β和IL-18。
1. 預激活后:NLRP3表達并結合于內質網和反式高爾基體網絡(TGN)的胞內膜,組裝為寡聚化雙環籠狀結構(PYD結構域包埋在內),此為無活性狀態;
2. 激活后:NLRP3激活劑誘導微管的聚合和乙酰化,反式高爾基體網絡囊泡與NLRP3分散,并被轉運至微管組織中心(MTOC),與NEK7結合形成預激活炎癥小體復合物(無Asc和半胱天冬酶-1前體);
3. 無活性的NLRP3寡聚籠狀結構通過結合乙酰化α-微管蛋白的HDAC6-動力蛋白復合物,沿微管從反式高爾基體網絡轉運至微管組織中心;
4. 反式高爾基體網絡囊泡通過微管釋放,使NLRP3重新定位至線粒體附近的細胞質,線粒體上的Asc和半胱天冬酶-1前體與NLRP3近距離接觸并相互作用,形成活性NLRP3炎癥小體復合物。
關鍵發現:α-微管蛋白的乙酰化是NLRP3激活的核心步驟,其機制為NLRP3激活引發線粒體功能障礙,進而導致乙酰化α-微管蛋白累積。
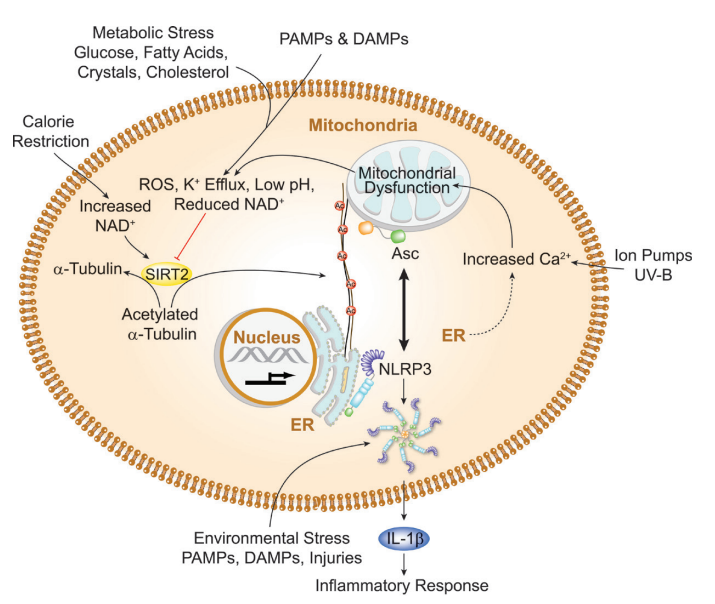
實驗圖:乙酰化α-微管蛋白的累積通過促進線粒體上的Asc與內質網上的NLRP3相互作用,推動炎癥小體的組裝和激活。
1. 小鼠全長NLRP3結構揭示的NLRP3籠狀結構調控通路激活,L. Andreeva等,《細胞》,2021年,第184卷,6299頁
2. 籠狀NLRP3調控炎癥小體活性,K. Schroder & R.C Coll,《細胞》,2021年,第184卷,6224頁
產品名稱 | 貨號 | 規格 | 來源 | 應用 |
抗乙酰化α-微管蛋白單克隆抗體(TEU318) | AG-20B-0068 | 100μg | 小鼠IgG1 | ICC、WB |
抗糖基化微管蛋白多克隆抗體(Gly-pep1) | AG-25B-0034 | 100μg | 兔 | ICC、IP、WB |
抗GTP化微管蛋白重組單克隆抗體(MB11) | AG-27B-0009 | 100μg | 人IgG2 λ | ICC |
抗β-微管蛋白(β-單谷氨酸化)多克隆抗體(IN115) | AG-25B-0039 | 50μg | 兔 | ICC、IHC、IP、WB |
抗多聚谷氨酸化修飾單克隆抗體(GT335) | AG-20B-0020 | 100μg | 小鼠IgG1k | 電鏡(EM)、ICC、IP、WB |
抗多聚谷氨酸化修飾單克隆抗體(GT335,生物素標記) | AG-20B-0020B | 100μg | 小鼠IgG1k | ICC、IP、WB |
抗多聚谷氨酸鏈(多聚E)多克隆抗體(IN105) | AG-25B-0030 | 50μg | 兔 | ICC、WB |
1. 秋水仙堿(Colchicine),貨號AG-CN2-0048,規格500mg/1g:微管抑制劑,抑制乙酰化α-微管蛋白介導的線粒體轉運,進而阻斷線粒體上的Asc與內質網上的NLRP3的相互作用(相關文獻:微管驅動的線粒體空間排布促進NLRP3炎癥小體激活,T. Misawa等,《自然免疫學》,2013年,第14卷,454頁);
2. 動力蛋白抑制劑:Dynasore(AG-CR1-0045);
3. F-肌動蛋白穩定劑:Jasplakinolide(AG-CN2-0037);
4. F-肌動蛋白解聚劑:Latrunculin A(AG-CN2-0027)、Latrunculin B(AG-CN2-0031)、細胞松弛素B(AG-CN2-0504)、葫蘆素E(AG-CN2-0474);
5. F-肌動蛋白抑制劑:Swinholide A(AG-CN2-0035);
6. 微管抑制劑:秋水仙胺(AG-CR1-3567)、伊里諾醌(AG-CN2-0038)、諾考達唑(AG-CR1-0019)、鬼臼毒素(AG-CN2-0049)、擬海松酸B(AG-CN2-0083);
7. 微管穩定劑:紫杉醇(AG-CN2-0045);
8. 微管抑制劑:帚曲霉素A(AG-CN2-0515)。
相關文獻:微管驅動的線粒體空間排布促進NLRP3炎癥小體的激活,T. Misawa等,《自然免疫學》,2013年,第14卷,454頁
產品名稱 | 作用靶點 | 貨號 |
動力蛋白抑制劑(Dynasore) | 動力蛋白抑制劑 | AG-CR1-0045 |
Jasplakinolide | F-肌動蛋白穩定劑 | AG-CN2-0037 |
拉春庫林A(Latrunculin A) | F-肌動蛋白解聚劑 | AG-CN2-0027 |
拉春庫林B(Latrunculin B) | F-肌動蛋白解聚劑 | AG-CN2-0031 |
斯維內酯A(Swinholide A) | F-肌動蛋白抑制劑 | AG-CN2-0035 |
細胞松弛素B(Cytochalasin B) | 肌動蛋白解聚劑 | AG-CN2-0504 |
葫蘆素E(Cucurbitacin E) | 肌動蛋白解聚劑 | AG-CN2-0474 |
秋水仙胺(Colcemid) | 微管抑制劑 | AG-CR1-3567 |
伊里諾醌(Ilimaquinone) | 微管抑制劑 | AG-CN2-0038 |
諾考達唑(Nocodazole) | 微管抑制劑 | AG-CR1-0019 |
紫杉醇(Paclitaxel) | 微管穩定劑 | AG-CN2-0045 |
磷霉素A(Phomopsin A) | 微管抑制劑 | AG-CN2-0515 |
鬼臼毒素(Podophyllotoxin) | 微管抑制劑 | AG-CN2-0049 |
偽綿馬酸B(Pseudolaric acid B) | 微管抑制劑 | AG-CN2-0083 |
經典炎癥小體 | 非經典炎癥小體 |
關鍵組分:吡喃蛋白、NLRP1b、NLRP3、NLRC4、AIM2、半胱天冬酶-1前體 | 關鍵組分:半胱天冬酶-11/4/5前體、脂多糖(LPS) |
激活過程:傳感器蛋白識別刺激后,與銜接蛋白Asc、半胱天冬酶-1前體組裝形成寡聚化活性炎癥小體復合物 | 激活過程:半胱天冬酶-11/4/5前體識別LPS后發生寡聚化并激活 |
下游效應:半胱天冬酶-1激活,切割IL-1β前體(p30)產生活性IL-1β,切割氣膜蛋白D(GSDMD)釋放N端片段 | 下游效應:激活的半胱天冬酶-11/4/5切割氣膜蛋白D(GSDMD)釋放N端片段 |
最終結果:IL-1β分泌至細胞外,氣膜蛋白D N端片段在細胞膜形成孔道,引發焦亡和炎癥反應 | 最終結果:氣膜蛋白D N端片段在細胞膜形成孔道,引發焦亡和炎癥反應 |
抗小鼠氣膜蛋白D(C端)多克隆抗體(IN110)
貨號:AG-25B-0036,規格:100微克
• 抗體來源:豚鼠
• 免疫原:重組小鼠氣膜蛋白D(C端)
• 適用實驗:酶聯免疫吸附測定(ELISA)、蛋白質印跡(WB)
• 特異性:識別小鼠氣膜蛋白D的全長及切割型C端結構域,與人氣膜蛋白D無交叉反應
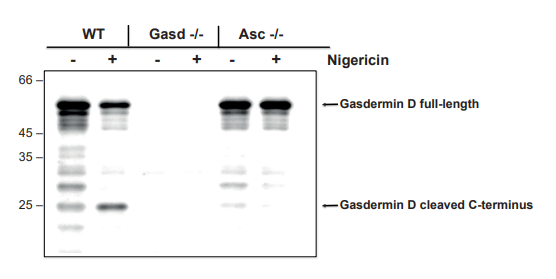
圖:采用抗小鼠氣膜蛋白D多克隆抗體(IN110,貨號AG-25B-0036),通過免疫印跡法檢測到小鼠氣膜蛋白D(全長及切割型p22片段)。
實驗方法:用50納克/毫升脂多糖(LPS,貨號AG-CU1-0001)處理野生型(WT)、氣膜蛋白D基因敲除(Gasd -/-)或Asc基因敲除(Asc -/-)小鼠的骨髓來源巨噬細胞(BMDMs)3小時,再用5微摩爾尼日利亞菌素(貨號AG-CN2-0020)處理2.5小時(或不處理),提取細胞提取物通過蛋白質印跡法分析氣膜蛋白D。將細胞提取物經還原條件下的SDS-PAGE分離,轉印至硝酸纖維素膜,與0.5微克/毫升的抗小鼠氣膜蛋白D多克隆抗體(IN110)孵育;加入1:5000稀釋的辣根過氧化物酶(HRP)偶聯抗豚鼠二抗后,通過化學發光檢測系統顯色。
圖片來源:德國弗萊堡大學醫學中心Olaf Gross教授提供
焦亡是一種炎癥性程序性細胞死亡,由病原體或宿主來源的細胞質擾動觸發。炎癥小體傳感器激活后,會激活半胱天冬酶-1及其他炎性半胱天冬酶,這些酶切割氣膜蛋白D和IL-1β/IL-18前體,最終引發焦亡和成熟細胞因子分泌。
焦亡以細胞死亡為代價,破壞胞內病原體的生存環境并釋放胞內成分,誘導鄰近細胞產生促炎癥反應。IL-1β是強效的促炎癥調控因子,可募集中性粒細胞、激活巨噬細胞并促進T細胞增殖。因此,焦亡和細胞因子分泌是炎癥小體信號通路下游的兩大核心機制,它們維持機體穩態、驅動固有免疫反應并塑造適應性免疫。
氣膜蛋白D(GSDMD)是焦亡的核心介導因子,包含功能性N端結構域和抑制性C端結構域。半胱天冬酶-1/11在氣膜蛋白D兩個結構域之間的連接區(人Asp275、小鼠Asp276)切割后,釋放的N端片段發生寡聚化,在宿主細胞膜形成孔道,最終導致細胞焦亡。此外,細胞表面死亡受體連接和寡聚化激活的半胱天冬酶-8,也可觸發氣膜蛋白D依賴的焦亡。
1. 炎癥小體信號通路下游焦亡與IL-1β分泌的解偶聯,Y. Li、Q. Jiang,《免疫學前沿》,2023年,第14卷,1128358頁
2. 焦亡的分子機制,J. Marisa等,《分子生物學方法》,2023年,第2641卷,1頁
3. 氣膜蛋白的“失控":氣膜蛋白家族在調控細胞穩態中的新作用,C.G. Weindel等,《細胞生物學趨勢》,2023年(在線優先出版)
4. 焦亡:一把兩刃劍——在炎癥性疾病和癌癥中的致病作用與治療潛力,Z. Liu等,《生命科學》,2023年,第318卷,121498頁
FACS:流式細胞術;FUNC:功能實驗;ICC:免疫細胞化學;IHC:免疫組化;IP:免疫沉淀;WB:蛋白質印跡;PF:無防腐劑;Hu:人;Ms:小鼠;Rt:大鼠;Rb:兔;Prm:靈長類
氣膜蛋白D N端片段在細胞膜形成孔道后,其C端片段的功能和去向仍不明確。本公司的小鼠氣膜蛋白D ELISA試劑盒(貨號AG-45B-0011)可檢測氣膜蛋白D的C端片段及全長蛋白,在焦亡細胞的上清液中可檢測到信號,表明C端片段會從細胞中釋放——可能是由于孔道存在被動釋放,也可能具有尚未明確的特定功能。
貨號:AG-45B-0011,規格:1塊96孔板
項目 | 說明 |
檢測靈敏度 | 14皮克/毫升 |
檢測范圍 | 15.6-1000皮克/毫升 |
適用樣本 | 細胞培養上清液、細胞裂解液 |
特異性 | 檢測細胞培養上清液和細胞提取物中小鼠氣膜蛋白D的全長及切割型C端片段,與人氣膜蛋白D無交叉反應 |
驗證實驗:
1. 對轉染脂多糖(LPS)的不同基因敲除小鼠品系的骨髓來源巨噬細胞(BMDMs)上清液進行檢測(見圖1),僅野生型(WT)和NLRP3基因敲除(NLRP3 KO)小鼠的上清液中檢測到氣膜蛋白D。
2. 對野生型和氣膜蛋白D基因敲除(Gasd KO)小鼠的骨髓來源巨噬細胞(BMDMs)細胞提取物(用Triton X-100緩沖液裂解)進行檢測(見圖2),僅野生型小鼠的細胞提取物中檢測到氣膜蛋白D。
貨號:AG-CR1-3698,規格:1毫克/5毫克
• 作用:氣膜蛋白D N端片段(GSDMD-N)誘導焦亡的抑制劑,可保護巨噬細胞免受GSDMD-N的細胞毒性,或保護小鼠免受致死性感染
• 分子式:C??H??O?
• 分子量:464.6
• CAS號:112648-68-7
相關文獻:脂質過氧化驅動氣膜蛋白D介導的焦亡在致死性混合菌膿毒癥中的作用,R. Kang等,《細胞宿主與微生物》,2018年,第24卷,97頁
貨號:AG-CR1-3705,規格:5毫克/25毫克
• 作用:氣膜蛋白D抑制劑,適用于小鼠研究,可直接結合氣膜蛋白D,抑制其N端片段寡聚化,進而阻斷孔道形成和焦亡
• 分子式:C??H??N?O?S
• 分子量:461.5
• CAS號:1360614-48-7
相關文獻:化學阻斷焦亡孔形成蛋白氣膜蛋白D可抑制炎癥性細胞死亡和膿毒癥,J.K. Rathkey等,《科學免疫學》,2018年,第3卷,eaat2738頁
半胱天冬酶-3/-7切割氣膜蛋白E(GSDME)后,其N端焦亡誘導結構域(GSDME-NT)從自身抑制的C端調控結構域中釋放,進而觸發細胞膜孔道形成和焦亡。人類氣膜蛋白E(GSDME)突變與遺傳性非綜合征性耳聾的發生相關。焦亡異常與多種人類疾病相關,包括感染、自身炎癥性疾病、神經退行性疾病和癌癥。
貨號:AG-45B-0024,規格:1塊96孔板
項目 | 說明 |
檢測靈敏度 | 50皮克/毫升 |
檢測范圍 | 0.625-4納克/毫升 |
適用樣本 | 細胞培養上清液、血漿、血清 |
特異性 | 檢測人氣膜蛋白E的C端結構域及全長蛋白;氣膜蛋白E切割和孔道形成后,其C端片段釋放到細胞外,可在血清/血漿中檢測到 |
Toll樣受體5(TLR5)可識別革蘭氏陽性菌和革蘭氏陰性菌的鞭毛蛋白。該受體激活后,通過銜接蛋白MyD88、TIRAP和TRIF介導的信號傳導,刺激腫瘤壞死因子-α(TNF-α)等促炎癥細胞因子的產生。
鞭毛蛋白是聚合形成細菌鞭毛絲的亞基蛋白,不僅可通過TLR5激活固有免疫系統,還能通過胞內NAIP5/NLRC4(IPAF)炎癥小體蛋白激活免疫反應。鞭毛蛋白激活NLRC4炎癥小體和TLR5,是機體對細菌感染啟動固有免疫反應的重要機制,在炎癥調控中起關鍵作用。NLRC4炎癥小體調控異常與膿毒癥、炎癥性腸病和某些類型癌癥的發生相關。
AdipoGen生命科學公司提供多種低內毒素、高純度的鞭毛蛋白產品,包括通路特異性突變體:
• 重組鞭毛蛋白(NLRC4突變體)(貨號AG-40B-0126):僅能被TLR5識別,無法被NLRC4識別
• 重組鞭毛蛋白(TLR5突變體)(貨號AG-40B-0127):僅能被NLRC4識別,無法被TLR5識別
產品名稱 | 貨號 | 規格 |
天然鞭毛蛋白 | AG-40B-0095 | 100微克 |
高純度天然鞭毛蛋白 | AG-40B-0025 | 10微克、3×10微克 |
重組鞭毛蛋白(帶組氨酸標簽) | AG-40B-0125 | 10微克、3×10微克 |
重組鞭毛蛋白(無標簽,高活性) | AG-40B-0243 | 50微克 |
重組鞭毛蛋白(NLRC4突變體) | AG-40B-0126 | 10微克、3×10微克 |
重組鞭毛蛋白(TLR5突變體) | AG-40B-0127 | 10微克、3×10微克 |
貨號:AG-CR1-3548,規格:10毫克/50毫克
• 體外實驗表明,m-3M3FBS可誘導線粒體損傷,并激活角質形成細胞中的NLRP10(與Asc和半胱天冬酶-1結合的功能性炎癥小體)
• NLRP10在腸上皮細胞中也可作為炎癥小體發揮作用,保護腸道免受自身炎癥損傷
相關文獻:
1. 線粒體損傷激活NLRP10炎癥小體,T. Prochnicki等,《自然免疫學》,2023年,第24卷,595頁
2. 上皮細胞Nlrp10炎癥小體介導對腸道自身炎癥的保護作用,D. Zheng等,《自然免疫學》,2023年,第24卷,585頁
AIM2炎癥小體抑制劑:蘇拉明鈉(Suramin sodium)
貨號:AG-CR1-3575V,規格:50毫克/250毫克/1克
• 蘇拉明是雙鏈DNA(dsDNA)誘導炎癥的有效抑制劑,可抑制多種雙鏈DNA結合蛋白,包括固有免疫通路中的cGAS/STING
• 新研究表明,蘇拉明可可逆性抑制小鼠和人的AIM2炎癥小體
相關文獻:一種優先靶向AIM2炎癥小體的DNA驅動炎癥抑制劑的發現,J.P. Green等,《交叉科學》,2023年,第26卷,106758頁
更多產品信息,請聯系中國區域代理商:上海起發實驗試劑有限公司
如果您對任意產品或其他品牌感興趣,可通過“在線留言"功能留下您的需求
(注:本文內容基于品牌公開資料及行業常規信息整理,具體以品牌信息文檔為準。)
電話
微信

 當前位置:
當前位置: